How does fatigue develop after exercise?
Please subscribe to my newsletter to read the future articles in this series!
Fatigue is a temporary reduction in exercise performance as a result of previous exercise. Since it is temporary, the mechanisms that cause fatigue during exercise dissipate, leading to a recovery of strength. However, at the same time as some fatigue mechanisms dissipate, others develop in a delayed fashion (in the same way as delayed onset muscle soreness). To understand fatigue in the hours and days following a workout, we therefore need to understand [A] how the fatigue mechanisms that arise during exercise dissipate, and [B] how the fatigue mechanisms that develop after exercise appear and then dissipate again. In a previous article, I focused solely on the dissipation of those fatigue mechanisms that actually develop during exercise, and in this article I will focus on the delayed fatigue mechanisms that only appear after exercise.
What mechanisms of fatigue develop during a bout of exercise?
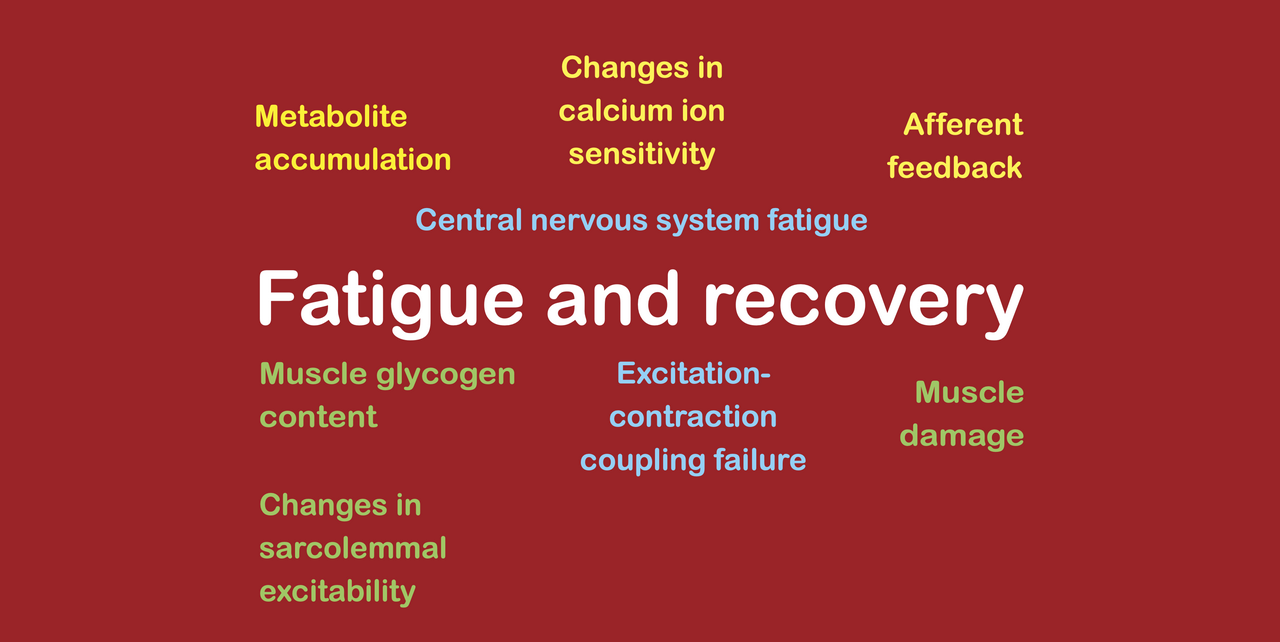
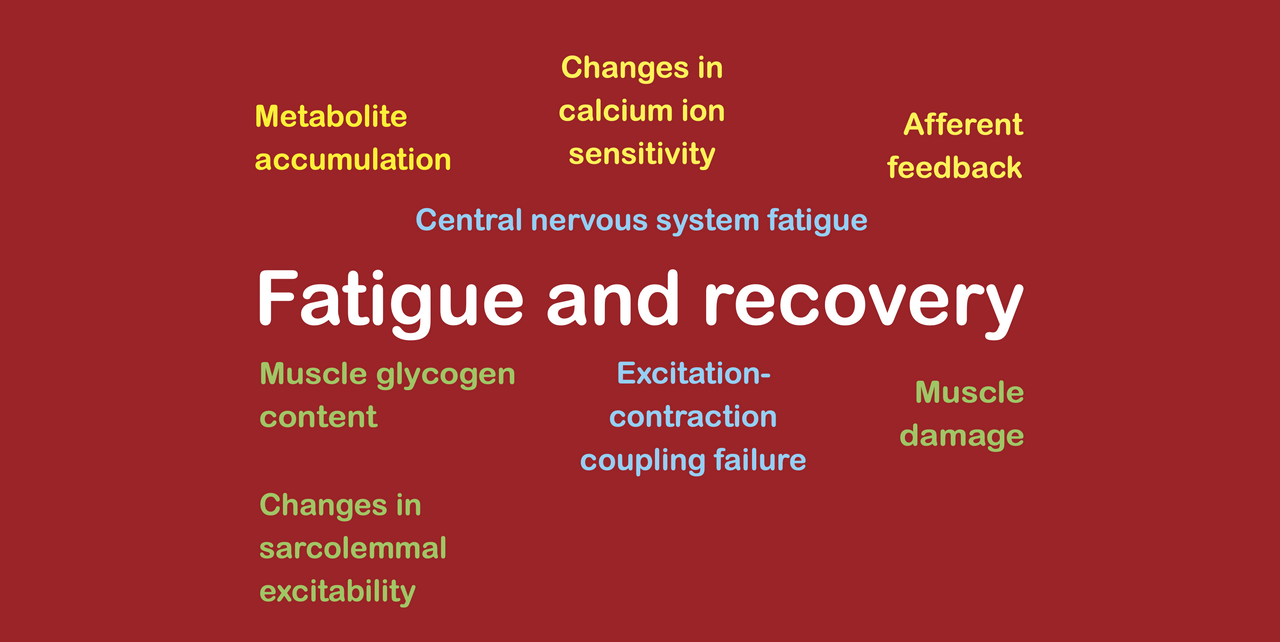
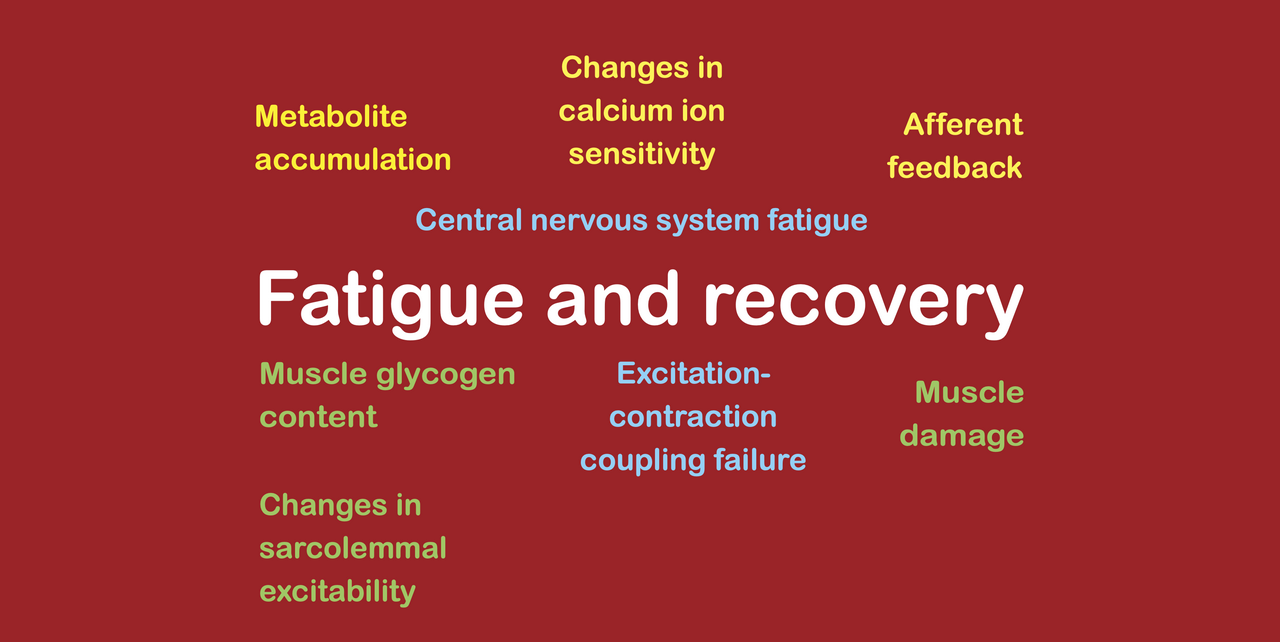
During exercise, fatigue can arise through several different fatigue mechanisms, including mechanisms. Considering all types of exercise (including strength training and other forms of anaerobic exercise as well as endurance exercise) we can identify several groups of fatigue mechanisms, which are as follows:
metabolite accumulation (largely strength training only);
glycogen depletion (largely aerobic exercise only);
calcium ion accumulation (including excitation-contraction coupling failure, losses of calcium ion sensitivity, and reductions in sarcolemmal excitability);
spinal CNS fatigue; and
supraspinal CNS fatigue (caused by afferent feedback due to metabolite accumulation or inflammatory mediators, and by inflammatory mediators entering the bloodstream).
Importantly, each of these fatigue mechanisms do not all dissipate at the same rate. Some fatigue mechanisms dissipate very quickly, while others dissipate much more slowly. Specifically, metabolite accumulation dissipates within approximately half an hour, glycogen depletion is reversed within a single day (although it likely causes this long-lasting fatigue through calcium ion-related mechanisms rather than directly by the absence of fuel), calcium ion-related fatigue can last for days if it is caused by excitation-contraction coupling failure, spinal CNS fatigue probably dissipates within a couple of minutes, and supraspinal CNS fatigue probably dissipates within half an hour (when it is caused by afferent feedback from metabolite accumulation) or a few hours (if caused by afferent feedback from inflammatory mediators). So the only long-lasting fatigue mechanism that occurs during exercise involves calcium ion accumulation in general and excitation-contraction coupling failure in particular.
What mechanisms of fatigue develop only after a bout of exercise?
#1. Muscle damage
Muscle damage is commonly believed to be generated during the bout of exercise itself. This is because it is assumed to occur as a result of the very high levels of mechanical tension that are present during eccentric contractions stretching muscle fibers until they break (leading to the “popping” of individual sarcomeres) as well as structural damage to the cytoskeleton and the muscle cell membrane. However, many studies have shown that muscle damage is not in fact present immediately after a bout of exercise that involves eccentric contractions. Rather, there is a delay of at least several hours before any signs of sarcomere rearrangement become apparent. How is this possible?
In fact, muscle damage is not actually caused by the high levels of mechanical tension that occur during eccentric contractions stretching muscle fibers until they break. Muscle fibers probably only physically break during repeated stretches if they are either [A] exposed to lengths that are way beyond those that can be achieved physiologically, or [B] excessively fatigued before being actively stretched. And these circumstances are more accurately described as muscle strain injury rather than muscle damage.
Rather, most muscle damage is actually caused by a biochemical effect, resulting from the influx and accumulation of calcium ions into the muscle fiber when muscle activation is sustained during a bout of exercise. When calcium ions are released out of the sarcoplasmic reticulum to stimulate the actin-myosin crossbridge binding process that causes muscle fibers to produce force, they are usually taken back up into the sarcoplasmic reticulum immediately afterwards. However, when contractions are repeated and muscle activation is sustained, calcium ions can accumulate inside the muscle fiber. If the mitochondria do not remove them quickly enough, the presence of these calcium ions then triggers a signaling response leading to the production of calpains (proteases) that degrade the insides of the cell. Researchers have shown that there is a pronounced increase in calpain levels immediately after a bout of muscle-damaging exercise, which is related to the sustained fatigue that is experienced.
While some commentators have assumed that the tendency for eccentric contractions to cause more muscle damage than concentric and isometric contractions is clear proof that mechanical tension is involved in producing muscle damage, this claim totally ignores the fact that eccentric contractions cause the opening of stretch-activated ion channels that allow the influx of additional calcium ions into the muscle fiber, causing a large stimulus for calpain production and therefore both muscle damage and excitation-contraction coupling failure. It is not necessary to resort to the high mechanical tension that is produced during eccentric contractions in order to explain how large amounts of muscle damage are produced.
Indeed, the process that causes muscle damage is clearly similar to the process that causes excitation-contraction coupling failure. In fact, it is identical, except that the calpains attack the minor triadic proteins that lie between the voltage sensor of the transverse tubules and the calcium ion store of the sarcoplasmic reticulum. Therefore, it is valuable to compare and contrast the development of excitation-contraction coupling failure with the development of muscle damage. In this respect, it is noteworthy that while excitation-contraction coupling failure clearly develops fairly rapidly during a bout of exercise (albeit not nearly as rapidly as metabolite accumulation during strength training), muscle damage does not become apparent until hours after a bout of exercise.
Excitation-contraction coupling failure occurs relatively quickly during a bout of exercise, partly because it can probably be temporarily created by other mechanisms and partly because it seems to require very little calpain activity for damage to occur to the minor proteins of this triadic junction such that excitation-contraction coupling failure occurs. In fact, this makes sense, because the triadic junction works by maintaining a close contact between the voltage sensor and the calcium ion store. Even a small disruption to the small, triadic proteins that hold the two structures adjacent to one another can result in the electrical signal that arrives at the voltage sensor failing to trigger the release of calcium ions into the cytoplasm from the calcium ion store. In contrast, muscle damage refers to wholesale restructuring of myofibrils and the cytoskeleton, which are much larger than the minor triadic proteins. Thus, the process of causing damage to them takes longer to play out, and it is not apparent until several hours after the workout has been completed.
#2. Supraspinal central nervous system (CNS) fatigue
While it is true that supraspinal CNS fatigue occurs during exercise, the nature of that supraspinal CNS fatigue is very transitory. Thus, we certainly cannot say that the supraspinal CNS fatigue that is produced during exercise contributes to the supraspinal CNS fatigue that persists several hours and days after a workout has been completed. Nevertheless, supraspinal CNS fatigue can be measured several days after exercise, which means that some other sources of supraspinal CNS fatigue must arise once the workout has finished.
During a strength training set, supraspinal CNS fatigue arises as a result of the afferent feedback from metabolites accumulating inside the muscle. The accumulating metabolites stimulate metaboreceptors that then send information back up afferent nerves to the brain, where it is registered as fatiguing or burning sensations. These perceptions of fatiguing or burning are then added to the perception of effort, which reduces the magnitude of the central motor command that can be attained at the maximal tolerable effort level. This in turn causes supraspinal CNS fatigue, which is an inability to produce the normally maximal level of central motor command and therefore the normally maximal level of motor unit recruitment, despite exerting a maximal effort. A similar phenomenon occurs during aerobic exercise, where supraspinal CNS fatigue arises as a result of the afferent feedback from inflammatory mediators accumulating inside the muscle (noting that these inflammatory mediators also enter the bloodstream and communicate with the brain through this system as well as by means of afferent feedback). Nevertheless, once the metabolites and the inflammatory mediators have dissipated, the supraspinal CNS fatigue similarly goes away. Therefore, this type of supraspinal CNS fatigue can only last for an hour or two after a bout of exercise, at the very most.
Nevertheless, the muscle damage response that occurs once exercise has been completed typically involves an inflammatory response. This inflammatory response is integral to the muscle damage and repair processes. Yet, it also causes supraspinal CNS fatigue in much the same way as the inflammatory mediators cause supraspinal CNS fatigue during aerobic exercise (as well as during long strength training workouts). While it has yet to be shown that these particular inflammatory mediators can cause supraspinal CNS fatigue by means of afferent feedback from the muscles, they have been found to cause supraspinal CNS fatigue by entering the bloodstream and causing increases in perceptions of fatigue and effort by acting directly on the brain. Therefore, the supraspinal CNS fatigue that is observed in the days after a strength training workout or after a bout of aerobic exercise is not caused by the same phenomena as the supraspinal CNS fatigue that was observed during the exercise bouts themselves.
What does this mean in practice?
Given that the only long-lasting fatigue mechanism that occurs during exercise is excitation-contraction coupling failure and given that the only mechanisms of fatigue that occur after exercise are muscle damage and supraspinal CNS fatigue, we can identify these three types of fatigue as being the sole causes of sustained fatigue in the days after a workout. This is important, since muscle damage and excitation-contraction coupling failure are both triggered by the same ultimate cause, which is the accumulation of calcium ions inside a muscle fiber as a result of a sustained period of muscle activation, and supraspinal CNS fatigue is itself caused by the inflammatory response to muscle damage. This means that there is actually only one ultimate cause of the sustained fatigue that is present in the days after a workout, and that is the accumulation of calcium ions during the workout as a result of the sustained period of muscle activation.
Consequently, within the context of a single contraction mode (eccentric, concentric, or isometric), we can now predict the amount of sustained fatigue that a single muscle fiber will experience after a workout based on the duration of time for which it is activated. Moreover, we can also predict the amount of fatigue that a whole muscle will experience after a workout based on the duration of time for which it is activated.
Unfortunately, these relationships between muscle activation time and muscle damage are not linear for either muscle fibers or whole muscles, because calcium ion accumulation seems to remain relatively low (likely while mitochondria are removing the ions) until a certain time point, after which it accelerates fairly quickly. Indeed, this is why proximity to muscular failure affects fatigue during strength training, because it is the final two reps of a set that seem to involve a large increase in excitation-contraction coupling failure. Also, the relationship between the duration of time for which a whole muscle is activated and the resulting fatigue will not be linear because slow twitch muscle fibers are less susceptible to calcium ion accumulation than fast twitch muscle fibers (since they have more mitochondria). Thus, very long workouts or bouts of aerobic exercise during which only slow twitch muscle fibers are active will not cause a huge amount of sustained fatigue, while even relatively short workouts or bouts of strength training during which fast twitch muscle fibers are active have the potential to cause far more sustained fatigue.
Nevertheless, even though the relationships are not linear, we can still say that the primary factor that causes fatigue after a workout is the duration of time for which muscle fibers and whole muscles are activated. This gives us a principle that we can use to minimize the sustained fatigue that occurs, since we can now aim to achieve a stimulus for whatever outcome is required (increased maximum strength, speed, muscle size) by maximizing the stimuli that cause adaptations suitable for those outcomes (increased ability to recruit motor units, increased coordination, increased muscle fiber diameter and length) while simultaneously reducing the duration of time for which muscle fibers are activated. For example, if we consider that there are only five or six stimulating reps per set to muscular failure, then we should be able to minimize fatigue by working with sets of 5–7 reps rather than by working with higher numbers of reps, where the added muscle activation time does not add any incremental benefit but causes much more sustained fatigue to accumulate.
What is the takeaway?
Fatigue is a temporary reduction in exercise performance as a result of previous exercise. During strength training exercise, fatigue is caused by metabolite accumulation, calcium ion accumulation, spinal CNS fatigue, and supraspinal CNS fatigue. During aerobic exercise, fatigue is caused by glycogen depletion, calcium ion accumulation, spinal CNS fatigue, and supraspinal CNS fatigue. After exercise, only excitation-contraction coupling failure (a type of fatigue caused by calcium ion accumulation) persists. However, muscle damage and supraspinal CNS fatigue caused by the inflammatory response subsequent to muscle damage arise in the hours and days following the exercise bout, such that the only three types of fatigue that are present in a sustained way are [1] excitation-contraction coupling failure, [2] muscle damage, and [3] supraspinal CNS fatigue. Moreover, since excitation-contraction coupling failure and muscle damage are caused by calcium ion accumulation subsequent to muscle activation, and since supraspinal CNS fatigue is caused by the inflammatory response to muscle damage, all three sustained fatigue mechanisms are ultimately caused by the exposure of muscle fibers to a sustained period of activation. Therefore, minimizing muscle activation duration while maximizing the key stimuli that cause adaptations is the key to optimizing strength training frequency.